










发酵神曲(Massa Medicata Fermentata, MMF)是一种中药,为辣蓼、青蒿、杏仁等药加入面粉或麸皮混合后,经发酵而形成的曲剂。临床上常用的MMF有两种形式,即生制MMF和炒制MMF。
根据中医理论,炒制能引起中药生物活性的变化。炒制MMF比生制MMF有更强的健脾作用。非挥发性和挥发性化合物的含量在翻炒后发生变化,pH值降低。与中性多糖相比,酸性多糖由于其结构特性(包括糖醛酸含量、分子量和单糖组成)而具有更好的生物活性。但是关于MMF中酸性多糖在炒制前后结构特征和生物活性变化的研究较少,导致MMF中活性物质作用应用有限。因此,在前期研究的基础上,进一步研究生制、炒制MMFs中酸性多糖的精确结构,将有助于厘清其生物活性的差异。
2024年1月,齐鲁理工大学在International Journal of Biological Macromolecules发表了”Structure and properties of acidic polysaccharides isolated from Massa Medicata Fermentata: Neuroprotective and antioxidant activity”的文章。作者通过柱层析分离纯化生制和炒制的MMFs中的酸性多糖,并利用分子量、单糖组成、核磁等现代方法比较它们的结构性质。阐述了神曲多糖在斑马鱼体内抗氧化和神经保护活性。确定了从MMF中提取的酸性多糖的构效关系。该研究为探讨MMF的炒制机理提供了参考,并为MMF及其炒制产物中活性物质的研究提供了理论依据。

SMMFAP:生制MMF的酸性部分;
CMMFAP:炒制MMF的酸性部分。
1.神曲多糖的理化结果及分子量分布
测定了神曲多糖的理化性质,结果表明SMMFAP和CMMFAP的总多糖及蛋白含量分别为71.86±0.21%和78.56±0.57%以及7.82±0.09%和7.13±0.57%。与之前的研究相比,生制MMF中酸性多糖的总多糖含量低于中性多糖,而蛋白质含量高于中性多糖。而炒制MM则是相反,可能炒的过程产生了这些影响。
SMMFAP和CMMFAP均为单峰。SMMFAP和CMMFAP的平均Mw分别为26.437和15.565 kDa(图1)。显然,在炒制过程中经过高温处理后,多糖的平均Mw下降,这可能是由于高温去除结合水导致多糖结构破坏和降解。
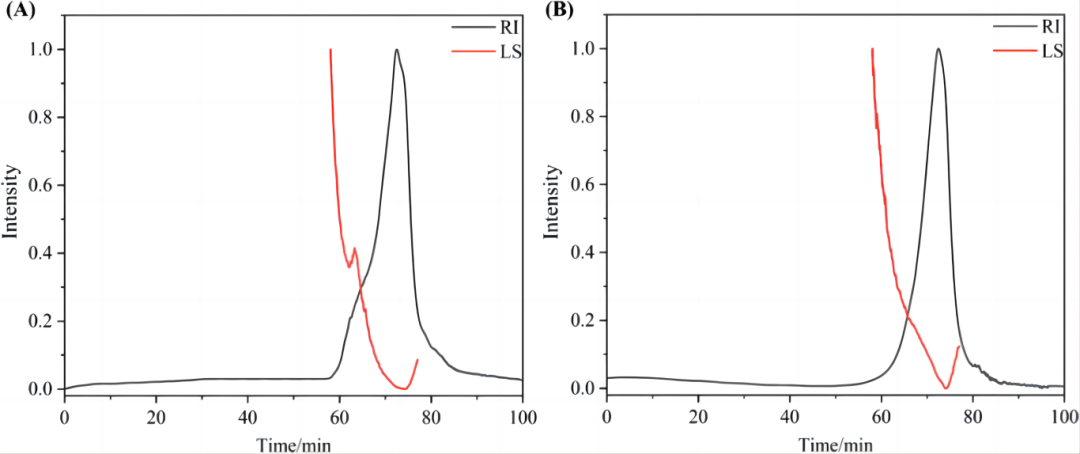
图1. SMMFAP (A) 和 CMMFAP (B)的平均分子量
2. 神曲多糖的单糖组成
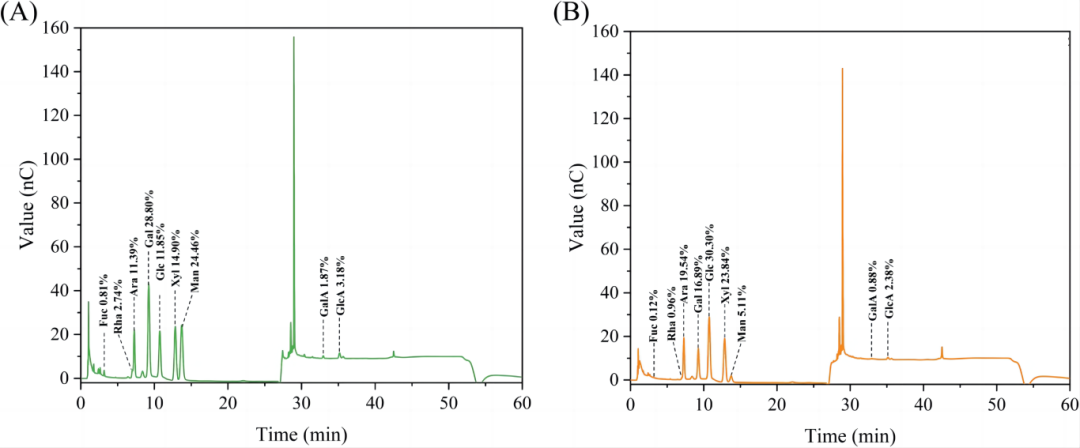
图2. 离子色谱的单糖结果,SMMFAP (A) 和 CMMFAP (B).
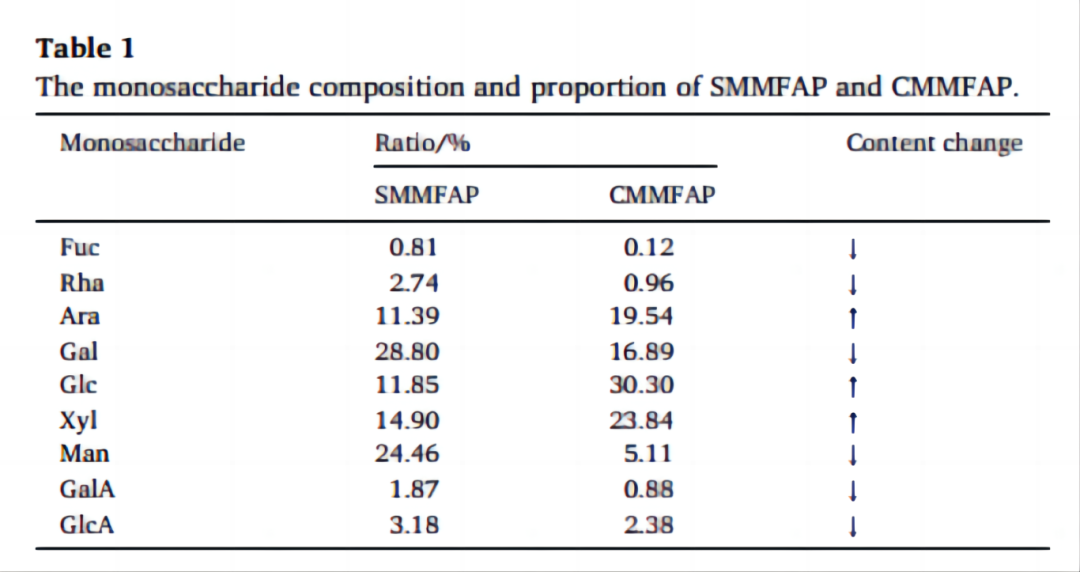
表1. 离子色谱的单糖结果,SMMFAP (A) 和 CMMFAP (B).
3.神曲多糖的红外结果
SMMFAP和CMMFAP的FTIR光谱(见图3),在3368 cm-1附近观察到的宽频带与-OH基团的伸缩振动吸收峰有关,在2921 cm-1附近是C-H的吸收峰。CMMFAP和SMMFAP的整体结果相似,例如1664 cm-1处的吸收带是羧基C—O的特征,表明存在醛酸。1407 cm-1处的吸收归因于GalA和GlcA的对称C—O拉伸等。
与CMMFAP相比,SMMFAP具有更高的β型糖苷键含量,根据单糖组成分析结果,这可归因于Gal和Man的相对比例降低,Ara的相对比例提高。
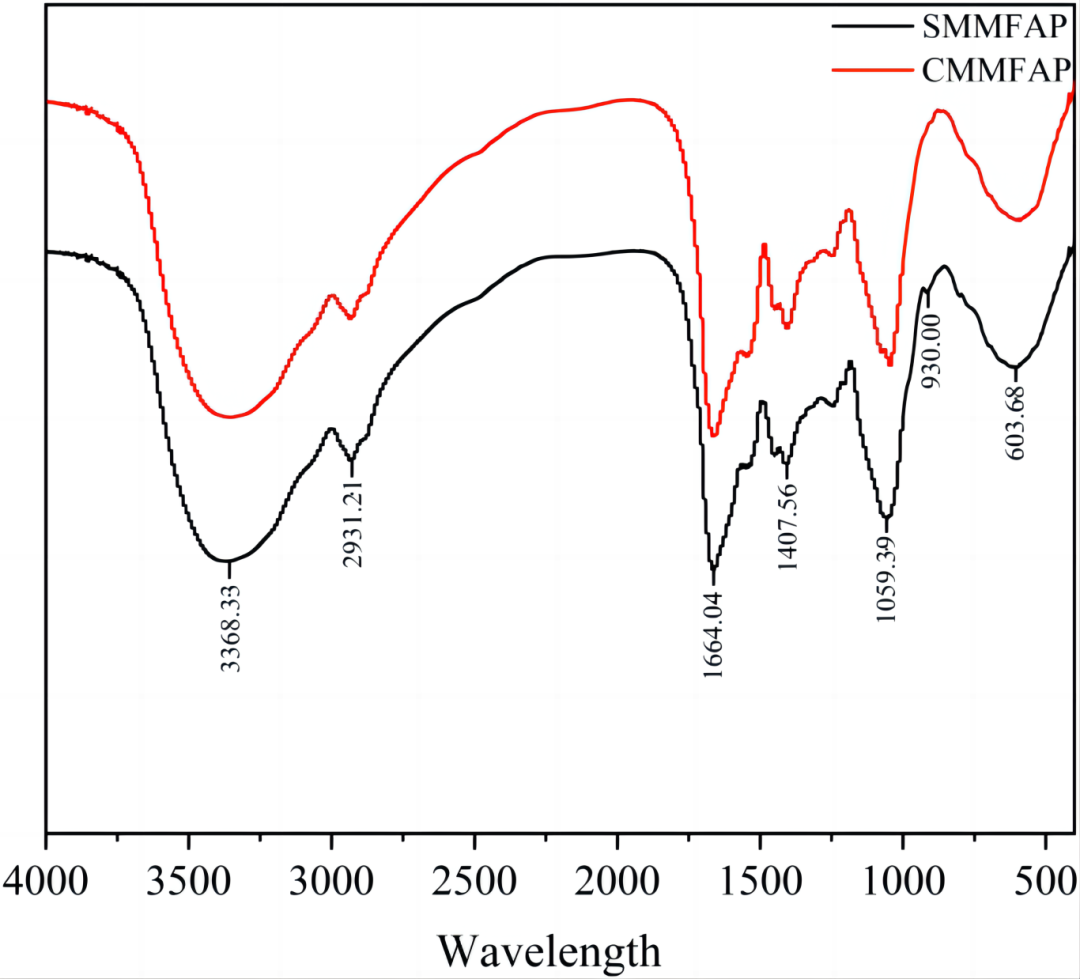
图3. SMMFAP (A) 和 CMMFAP (B).的红外结果
4.神曲多糖的甲基化结果
为了研究糖苷键的类型,对SMMFAP和CMMFAP键合结构解析 (图4)。结果表明SMMFAP和CMMFAP都是由不同的键合结构组成的,并且这些多糖的结构相对复杂。
甲基化分析结果表明,SMMFAP和CMMFAP都是由末端加链糖组成,主要构成有所不同。
与CMMFAP相比,SMMFAP的糖苷键类型更为复杂,说明在高温炒制处理下,一些糖苷键断裂并重新连接。另外,经过炒制,生成了新的糖苷键类型(1,4,6- glcp),这可能是由于糖苷键在高温处理下首先断裂,然后一些新的糖苷键发生氧化还原反应。这些酸性多糖的糖苷连接类型与MMF中的中性多糖无关,这表明酸性多糖不是MMF在炒制过程中中性多糖的反应产物。
图4. SMMFAP (A) 和 CMMFAP (B).的甲基化结果
5.神曲多糖的核磁分析
对神曲多糖的1D (1H和13C)和2D (COSY, HSQC和HMBC) NMR进行了分析(图5-6)。在1H谱图中,许多质子共振信号分布在δ 3.0-5.5 ppm。其中,己糖的端粒质子信号(H1)主要集中在δ 4.5 ~ 5.5 ppm。
H谱中,在δ4.53、5.20、5.21、5.17、5.03、4.50、4.44、5.01、4.56、5.42、4.41、4.48、5.30、4.52和4.94 ppm的浓度下,观察到15个不同的异头质子信号,分别对应于糖残基A、B、C、D、E、F、G、H、I、J、K、L、M和N。再结合二维核磁信息,对SMMFAP和 CMMFAP分别进行归属。
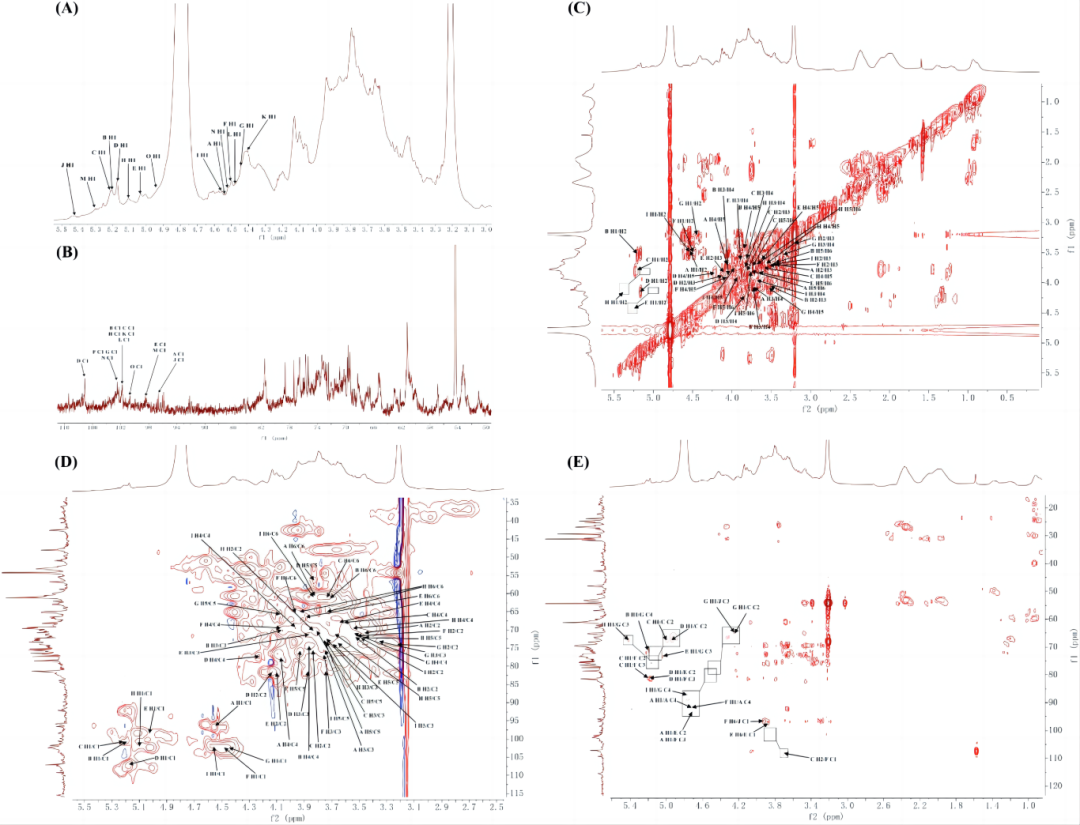
图5. SMMFAP的核磁谱图: 1H (A), 13C (B), COSY (C), HSQC (D) and HMBC (E).
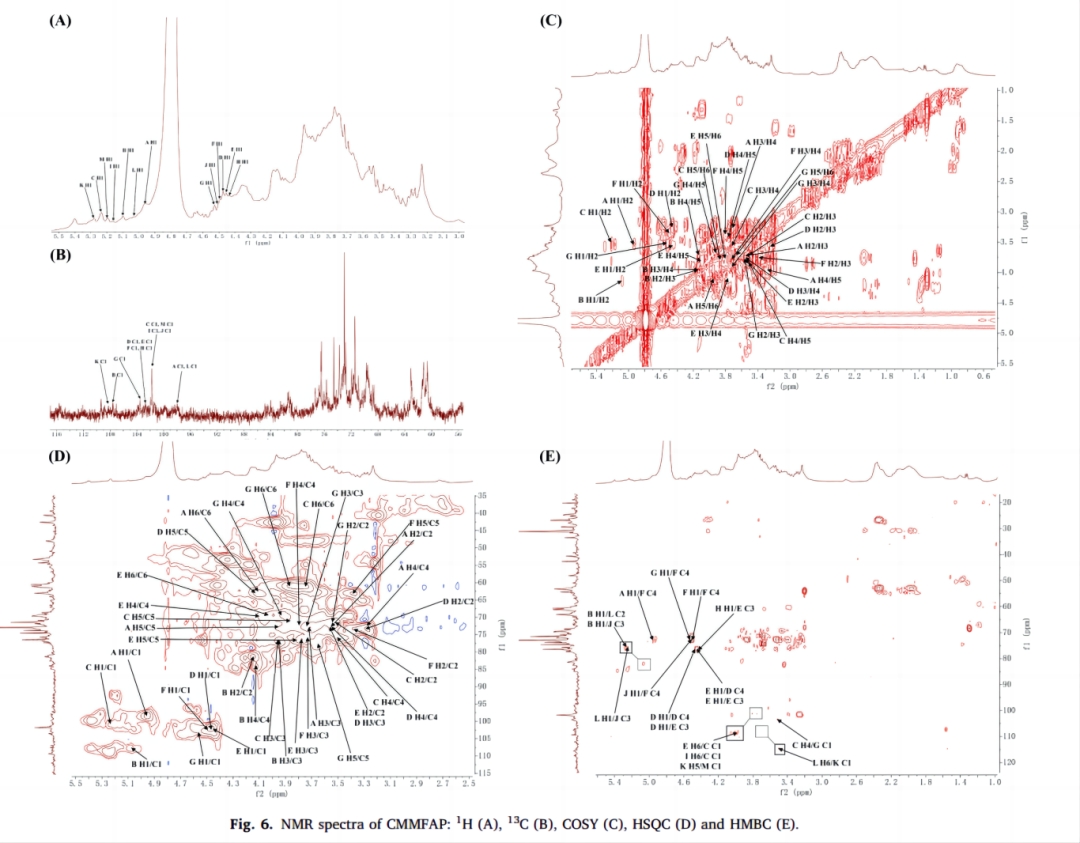
图6. CMMFAP的核磁谱图: 1H (A), 13C (B), COSY (C), HSQC (D) and HMBC (E).
6.神曲多糖的电镜结果
观察SMMFAP和CMMFAP的表面形貌。两种多糖的表面形态有显著差异。如图7所示,SMMFAP呈不规则多孔结构,而CMMFAP则由零散片状组成。这些结果表明SMMFAP可能具有更高的分支程度和更广泛的网络结构。CMMFAP的破碎结构可能会降低其溶解度和润湿性。

图7. SMMFAP (A) 和 CMMFAP (B).的电镜结果
7.体内神经保护活性
在测试最大耐受浓度的实验中,观察到200 μg/mL剂量组的斑马鱼均运动缓慢,死亡较多。在此基础上,选择25、50和100 μg/mL的浓度进行进一步研究。
如图8A、C所示,空白组多巴胺神经元区域长度(KB)明显长于模型组(MX),说明神经损伤模型建立成功。在多糖处理组,多巴胺神经元区长度随着多糖浓度的增加而逐渐增加,并且SMMFAP和CMMFAP均具有浓度依赖性的神经保护作用。
此外,当浓度为100 μg/mL时,CMMFAP的神经保护活性优于SMMFAP(图8D)。这些结果表明,CMMFAP的神经保护能力强于SMMFAP,使其成为治疗神经损伤的有希望的候选者。
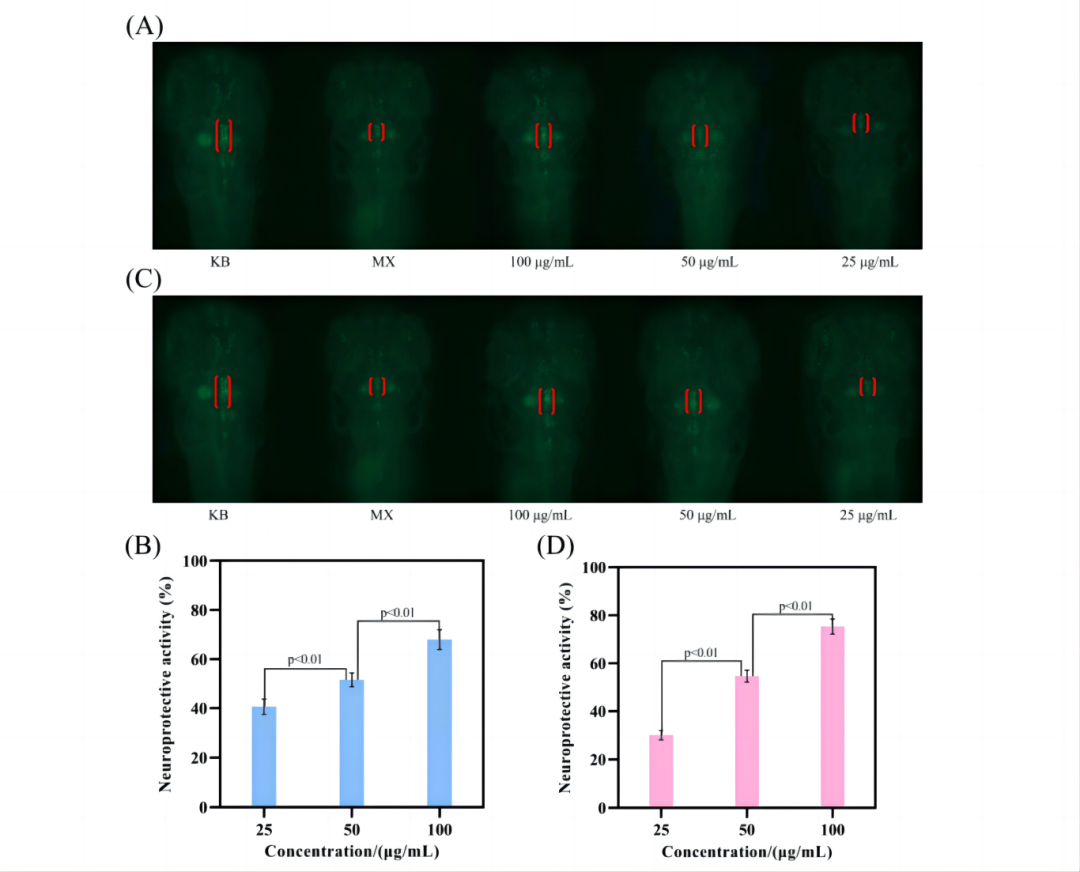
图8. 神经保护性研究。SMMFAP(AB), CMMFAP(CD)
8.体内抗氧化活性
如图所示,空白组有清晰的荧光点,而模型组只有少量荧光点,说明氧化损伤模型建立成功。在多糖组中,荧光点数目随多糖浓度的增加而增加,结果表明,SMMFAP和CMMFAP都具有抗氧化活性,并表现出强烈的浓度依赖性,且SMMFAP的抗氧化能力更强。
此外,神经保护活性的变化趋势与抗氧化活性并不一致,这表明多糖在体内的神经保护活性并不仅仅归因于其抗氧化能力,这与以往的研究结果不同。
这些结果进一步证明了这些酸性多糖的神经保护作用机制的复杂性。
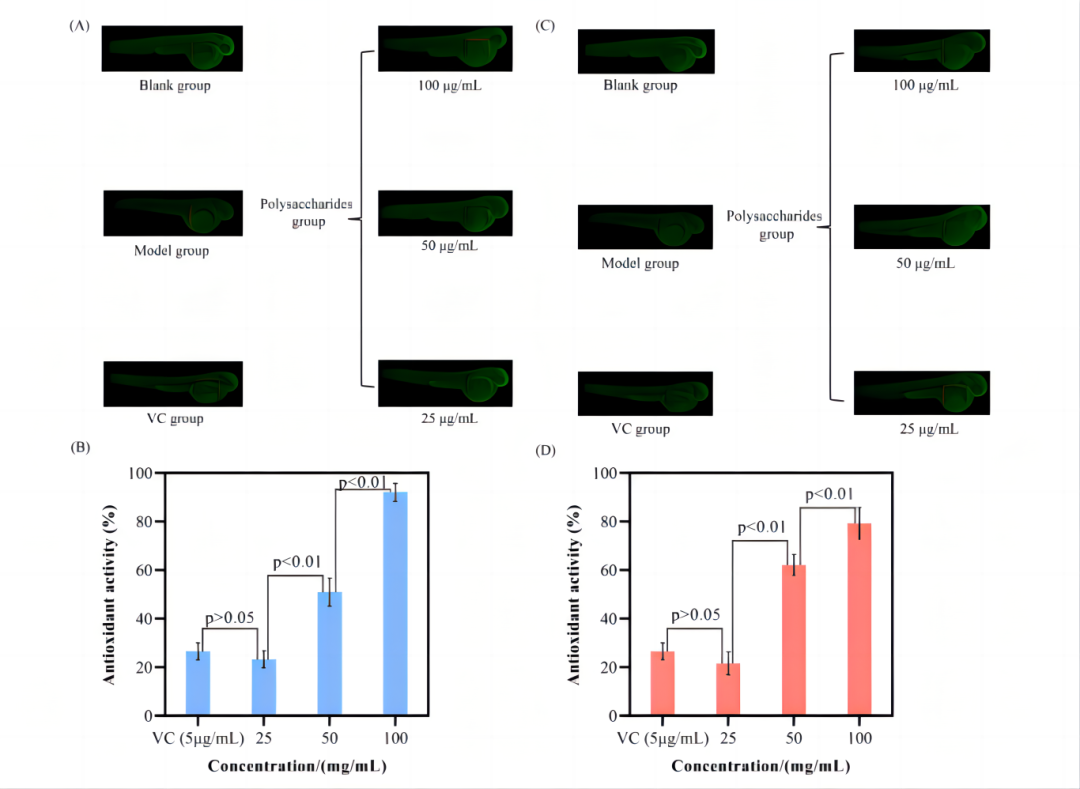
图9. 体内抗氧化研究。SMMFAP(AB), CMMFAP(CD)
9.网络药理学分析
本项研究使用网络药理学分析鉴定了180个靶点。共发现与神经损伤相关的靶点566个,与氧化应激相关的靶点646个。同时使用Venny平台筛选了17个潜在靶点。绿色,紫色和粉红色的节点分别代表酸性多糖潜在的目标和途径。从网络上发现,这些酸性多糖的神经保护作用机制是复杂的,并且这些酸性多糖表现出协同作用。通过调控PI3K-Akt和NF-kappa B信号通路直接缓解神经损伤,并可通过HIF-1信号通路调节氧化应激间接治疗神经损伤。
10.神曲多糖的构效关系
多糖的生物活性与其单糖组成、平均分子量和化学结构显著相关。
总体而言SMMFAP在体内的抗氧化活性强于CMMFAP。一是SMMFAP具有较高的平均Mw,二是其葡萄糖醛酸含量更高,这可能允许SMMFAP提供更多的氢与自由基反应并将其转化为更稳定的产物,三是其不规则的多孔结构可以储存自由基,具有良好的抗氧化活性。
但是在神经保护活性方面,CMMFAP比SMMFAP作用更强,这可能是由于其平均Mw比更低而且Glc含量更多。总的来说,初级结构,包括单糖组成、平均分子量和表面形态,可以影响生物活性。
综上所述,分别从生制和炒制两种MMF中分离纯化了两种水溶性较好的酸性多糖(SMMFAP和CMMFAP)。SMMFAP的平均分子量高于CMMFAP,而CMMFAP的Ara、Glc和Xyl含量高于SMMFAP。此外,多糖中的一些糖苷键在翻炒后断裂并重新连接,如1,4--glcp变成1,4,6- glcp。生物活性评价结果表明,CMMFP的体内神经保护活性优于SMMFP,而SMMFP的体内抗氧化活性优于CMMFP。
这些结果表明,酸性多糖可能是导致生制、炒制MMF疗效不同的活性物质。值得注意的是,SMMFAP良好的抗氧化活性和CMMFAP良好的神经保护活性可能为MMF多糖在人体保健品中的进一步开发提供有价值的信息。尽管这些多糖具有多种潜在的应用前景,但其结构与生物活性之间的详细关系以及这些酸性多糖生物活性的机制仍有待进一步研究。
文献解读:
文献解读 | 多组学和多模式脑电图数据的融合有助于神经认知障碍的个性化诊断

排版:野凌
审核:三黍生物企宣

